Что образуют митохондрии. Митохондрии
О СЛОЖНОМ ПРОСТЫМ ЯЗЫКОМ.
Тема эта сложная и комплексная, затрагивающая сразу же огромное количество биохимических процессов происходящих в нашем организме. Но давайте все таки попробуем разобраться, что же такое митохондрии и как они работают.
И так, митохондрии это одна из самых важных составляющих живой клетки. Если говорить простым языком то можно сказать, что это энергетическая станция клетки. Их деятельность основана на окисление органических соединений и генерации электрического потенциала (энергии освободившейся при распаде молекулы АТФ) для осуществления мышечного сокращения.
Все мы знаем, что работа нашего организма происходит в строгом соответствии с первым законом термодинамики. Энергия не создается в нашем организме, а лишь превращается. Организм только выбирает форму трансформации энергии, не производя ее, от химической к механической и тепловой. Основным источником всей энергии на планете Земля является Солнце. Приходя к нам в форме света, энергия поглощается хлорофиллом растений, там она возбуждает электрон атома водорода и таким образом дает энергию живой материи.
Своей жизнью мы обязаны энергии маленького электрона.
Работа митохондрии заключается в ступенчатом переносе энергии электрона водорода между атомами металлов, присутствующих в группах белковых комплексов дыхательной цепи (электронно-транспортной цепи белков), где каждый последующий комплекс обладает более высоким сродством к электрону притягивая его, чем предыдущий, до тех пор, пока электрон не соединиться с молекулярным кислородом, обладающим наибольшим сродством к электрону.
Каждый раз при передачи электрона по цепи высвобождается энергия которая аккумулируется в виде электрохимического градиента и затем реализовывается в виде мышечного сокращения и выделения тепла.
Серия окислительных процессов в митохондрии позволяющая перенести энергетический потенциал электрона называется «внутриклеточным дыханием» или часто «дыхательной цепью», так как электрон по цепочки передается от атома к атому до тех пор пока не достигнет своей конечной цели атома кислорода.
Митохондриям нужен кислород для переноса энергии в процессе окисления.
Митохондрии потребляют до 80% кислорода который мы вдыхаем.
Митохондрия представляет из себя постоянную структуру клетки, расположенную в ее цитоплазме. Размер митохондрии обычно составляет от 0,5 до 1 мкм в диаметре. По форме она имеет зернистую структуру и может занимать до 20% объема клетки. Такая постоянная органическая структура клетки называется органелла. К органеллам относятся и миофибриллы – сократительные единицы мышечной клетки; и ядро клетки это тоже органелла. Вообще, любая постоянная структура клетки является органоидом-органеллой.
Открыл митохондрии и впервые описал немецкий анатом и гистолог Рихард Альтман в 1894 году, а название этой органелле дал другой немецкий гистолог К. Бенд в 1897 году. Но только в 1920 году, опять же немецкий биохимик Отто Вагбург, доказал, что с митохондриями связаны процессы клеточного дыхания.
Существует теория, согласно которой митохондрии появились в результате захвата примитивными клетками, клетками которые сами не могли использовать кислород для генерации энергии, бактерий протогенотов, которые могли это делать. Именно потому, что митохондрия ранее представляла из себя отдельный живой организм она и по сей день обладает собственным ДНК.
Митохондрии ранее представляли из себя самостоятельный живой организм.
В ходе эволюции прогеноты предали множество своих генов сформировавшемуся, благодаря повысившейся энергоэффективности, ядру и перестали быть самостоятельными организмами. Митохондрии присутствуют во всех клетках. Даже в сперматозоиде есть митохондрии.
Именно благодаря им приводится в движение хвостик сперматозоида осуществляющий его движение. Но особенно много митахондрий в тех местах, где необходима энергия для любых жизненных процессов. И это конечно прежде всего мышечные клетки.
В мышечных клетках митохондрии могут объединяться в группы гигантских разветвленных митохондрий, связанных друг с другом с помощью межмитохондриальных контактов, в которых они создают согласованную работающую кооперативную систему. Пространство в такой зоне имеет повышенную электронную плотность. Новые митохондрии образуются путем простого деления предыдущих органелл. Наиболее «простой» и доступный всем клеткам механизм энергетического обеспечения чаще всего называют общим понятием гликолиз.
Это процесс последовательного разложения глюкозы до пировиноградной кислоты. Если этот процесс происходит без участия молекулярного кислорода или с недостаточным его присутствием, то он называется анаэробный гликолиз. При этом глюкоза расщепляется не до конечных продуктов, а до молочной и пировиноградной кислоты которая далее претерпевает дальнейшие превращения в ходе брожения. Поэтому высвобождающейся энергии бывает меньше, но и скорость получения энергии быстрее. В результате анаэробного гликолиза из одной молекулы глюкозы клетка получает 2 молекулы АТФ и 2 молекулы молочной кислоты. Такой «базовый» энергетический процесс может протекать внутри любой клетки без участия митохондрий.
В присутствии молекулярного кислорода внутри митохондрий осуществляется аэробный гликолиз в рамках «дыхательной цепи». Пировиноградная кислота в аэробных условиях вовлекается в цикл трикарбоновых кислот или цикл Кребса. В результате этого многостадийного процесса из одной молекулы глюкозы образуется 36 молекул АТФ. Сравнение энергетического баланса клетки, имеющей развитые митохондрии и клетки, где они не развиты показывает (при достаточном количестве кислорода) различие в полноте использования энергии глюкозы внутри клетки почти в 20 раз!
У человека, волокна скелетных мышц можно условно разделить на три типа исходя из механических и метаболических свойств: — медленные окислительные; — быстрые гликолитические; — быстрые окислительно-гликолитические.

Быстрые мышечные волокна предназначены для выполнения быстрой и тяжелой работы. Для своего сокращения они используют в основном быстрые источники энергии, а именно криатинфосфот и анаэробный гликолиз. Содержание митохондрий в таких типах волокон значительно меньше чем в медленных мышечных волокнах.
Медленные мышечные волокна выполняют медленные сокращения, но способны работать длительное время. В качестве энергии они используют аэробный гликолиз и синтез энергии из жиров. Это дает гораздо больше энергии чем анаэробный гликолиз, но требует в замен больше времени, так как цепочка деградации глюкозы более сложная и требует присутствия кислорода, транспортировка которого к месту преобразования энергии тоже занимает время. Медленные мышечные волокна называют красными из-за миоглобина – белка, ответственный за доставку кислорода внутрь волокна. Медленные мышечные волокна содержат значительное количество митохондрий.
Возникает вопрос, каким образом и с помощью каких упражнений можно развить в мышечных клетках разветвленную сеть митохондрий? Существуют различные теории и методики тренировок и о них в материале по ссылке.
Характеристика, роль и строение митохондрий
Функции митохондрий как органелл аэробных эукариотических клеток – синтез молекул АТФ (аденозинтрифосфата) из АДФ. Поскольку АТФ является универсальным источником энергии для всех процессов в клетке, идущих с потреблением энергии, то говорят, что митохондрии выполняют функцию энергообеспечения, или энергообразования .
Из цитоплазмы в митохондрии поступают промежуточные продукты окисления органических веществ, кислород, АДФ, фосфорная кислота. Обратно выделяются углекислый газ, вода и молекулы АТФ.
Молекулы АТФ образуются не только в митохондриях. Небольшое их количество синтезируется в цитоплазме в процессе гликолиза, который наблюдается во всех клетках живого. В результате гликолиза молекула глюкозы разлагается на две молекулы пирувата. У аэробных прокариот далее он окисляется в присутствии кислорода на впячиваниях цитоплазматической мембраны. У эукариот же пируват поступает в митохондрии.
Здесь пируват, отдает свою ацетильную группу, содержащую два атома углерода, коферменту А. При этом выделяется первая молекула CO2. Кофермент А превращается в ацетил-кофермент-А (ацетил-КоА).
Ацетил-КоА получается не только из пирувата, но и жирных кислот, а также аминокислот. Так что не важно, какое исходное органическое вещество будет «сжигаться» в митохондриях для выработки энергии. Их функционирование в любом случае универсально.
В матриксе митохондрий ацетил-КоА вступает в цикл Кребса , или цикл трикарбоновых кислот, где ацетильная группа окисляется и разлагается до еще двух молекул CO2. Ее атомы водорода присоединяются к коферментам НАД+ и ФАД+, с образованием их восстановленных форм - НАД · H + Н+ и ФАД · H + Н+. Именно их последующее окисление приведет к синтезу АТФ.
Хотя в цикле Кребса кислород не используется, при его отсутствии митохондрия перестает выполнять свою функцию уже на этом этапе, так как накапливаются продукты реакции.
На кристах митохондрий происходит разделение электронов и протонов водорода. Электроны от НАД и ФАД передаются по мембране через цепь ферментов и коферментов, которую называют дыхательной цепью . Протоны же в начале пути перебрасываются в межмембранное пространство, на внешнюю сторону крист.
Электроны в конечном итоге передаются на молекулу кислорода, она превращается в отрицательно заряженный ион. Между внешней и внутренней сторонами крист создается электрический потенциал, так как одна заряжена положительно, а другая – отрицательно. Когда достигается критическое значение H+ устремляются через каналы АТФ-синтетазы и другие ферменты на внутреннюю сторону, где соединяются с O2- с образованием воды.
АТФ-синтетаза – это фермент, синтезирующий АТФ. В митохондриях он встроен в мембрану крист и использует энергию перемещающихся протонов для фосфорилирования АДФ.
Цикл Кребса и дыхательная цепь - это сложные многоступенчатые процессы, обеспечивающиеся целым рядом ферментов и коферментов. Каждый требует отдельного рассмотрения. В общих чертах же функции митохондрий сводятся к синтезу ацетил-КоА, использованию атомов водорода ацетильной группы для восстановления НАД и ФАД, раздельному переносу электронов и протонов водорода на кислород, использованию энергии электрохимического градиента протонов для синтеза АТФ.
Связанные статьи:Строение митохондрии, Этапы энергетического обмена
Митохондрия – это двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ – источника энергии для жизнедеятельности клетки.
Количество митохондрий в клетках не постоянно, в среднем от нескольких единиц до нескольких тысяч. Там, где процессы синтеза идут интенсивно, их больше. Также варьирует размер митохондрий и их форма (округлые, вытянутые, спиральные, чашевидные и др.). Чаще имеют округлую вытянутую форму, диаметром до 1 микрометра и длиной до 10 мкм. Могут перемещаться в клетке с током цитоплазмы или оставаться в одном положении. Перемещаются к местам, где больше всего требуется выработка энергии.
Согласно гипотезе симбиогенезамитохондрии произошли от аэробных бактерий, внедрившихся в другую прокариотическую клетку . Эти бактерии начали снабжать клетку дополнительным количеством молекул АТФ, а получать от нее питательные вещества. В процессе эволюции они потеряли автономность, передав часть своих генов в ядро и став таким образом клеточной органеллой.
В клетках новые митохондрии появляются в основном путем деления ранее существующих, т. е. они не синтезируются заново, что напоминает процесс размножения и говорит в пользу симбиогенеза.
Строение и функции митохондрии
Митохондрия состоит из
двух мембран - внешней и внутренней ,
межмембранного пространства ,
внутреннего содержимого - матрикса ,
крист , представляющих собой выросты в матрикс внутренней мембраны,
собственной белок-синтезирующей системы: ДНК, рибосом, РНК ,
белков и их комплексов, в том числе большого количества ферментов и коферментов,
других молекул и гранул различных веществ, находящихся в матриксе.

Внешняя и внутренняя мембраны выполняют разные функции, поэтому различается их химический состав. Расстояние между мембранами составляет до 10 нм. Внешняя мембрана митохондрий по строению схожа с плазмалеммой, окружающей клетку, и выполняет в основном барьерную функцию, отграничивая содержимое органоида от цитоплазмы. Через нее проникают мелкие молекулы, транспорт крупных избирателен. В некоторых местах внешняя мембрана соединена с ЭПС, каналы которой открываются в митохондрию.
На внутренней мембране, в основном ее выростах - кристах, располагаются ферменты, образуя мультиферментативные системы. Поэтому по химическому составу здесь преобладают белки, а не липиды. Количество крист варьирует в зависимости от интенсивности процессов. Так в митохондриях мышц их очень много.
В некоторых местах внешняя и внутренняя мембрана соединяются между собой.
У митохондрий, также как у хлоропластов, есть своя белоксинтезирующая система - ДНК, РНК и рибосомы. Генетический аппарат представляет собой кольцевую молекулу – нуклеоид, как у бактерий. Рибосомы митохондрий растений схожи с бактериальными, у животных митохондриальные рибосомы мельче не только цитоплазматических, но и бактериальных. Часть необходимых белков митохондрии синтезируют сами, другую часть получают из цитоплазмы, так как эти белки кодируются ядерными генами.
Главная функция митохондрий - снабжать клетку энергией, которая путем многочисленных ферментативных реакций извлекается из органических соединений и запасается в АТФ. Часть реакций идет с участием кислорода, в других выделяется углекислый газ. Реакции идут как в матриксе (цикл Кребса), так и на кристах (окислительное фосфорилирование).
Следует иметь в виду, что в клетках АТФ синтезируется не только в митохондриях, но и в цитоплазме в процессе гликолиза. Однако эффективность этих реакций невысока. Особенность функции митохондрий в том, что в них протекают реакции не только бескислородного окисления, но и кислородный этап энергетического обмена.
Другими словами, функция митохондрий – активное участие в клеточном дыхании, к которому относят множество реакций окисления органических веществ, переноса протонов водорода и электронов, идущих с выделением энергии, которая аккумулируется в АТФ.
Ферменты митохондрий
Ферменты транслоказы внутренней мембраны митохондрий осуществляют активный транспорт АДФ и АТФ.
В структуре крист выделяют элементарные частицы, состоящие из головки, ножки и основания. На головках, состоящих из фермента АТФазы , происходит синтез АТФ. АТФаза обеспечивает сопряжение фосфорилирования АДФ с реакциями дыхательной цепи.
Компоненты дыхательной цепи находятся в основании элементарных частиц в толще мембраны.
В матриксе находится большая часть ферментов цикла Кребса и окисления жирных кислот.
В результате активности электротранспортной дыхательной цепи ионы водорода поступают в нее из матрикса, а высвобождаются на наружной стороне внутренней мембраны. Это осуществляют определенные мембранные ферменты.
Митохондрия
Разница в концентрации ионов водорода по разные стороны мембраны приводит к возникновению градиента pH.
Энергию для поддержания градиента поставляет перенос электронов по дыхательной цепи. Иначе ионы водорода диффундировали бы обратно.
Энергия градиента pH используется для синтеза АТФ из АДФ:
АДФ + Ф = АТФ + H2O (реакция обратима)
Образующаяся вода ферментативно удаляется. Это, наряду с другими факторами, облегчает протекание реакции слева направо.
Митохондрии
Пластиды и митохондрии растительной клетки: структура, функции, особенности строения в связи с биологическими функциями.
Митохондрии растительной клетки. Их структура и функции
Форма − округлые или гантелевидные тельца.
Размеры − длина 1-5 мкм, диаметром 0,4-0,5 мкм.
Количество в клетке − от десятков до 5 000.
Структура . Состоят в основном из белка (60-65 %) и липидов (30 %). Это двухмембранные органоиды. Толщина наружной и внутренней мембран − 5-6 нм каждая. Перимитохондриальное пространство (промежуток между мемранами) заполнено жидкостью типа сыворотки. Внутренняя мембрана образует различной формы складки − кристы . На внутренней поверхности внутренней мембраны расположены грибовидные частицы − оксисомы, содержащие окислительные ферменты. Внутреннее содержимое митохондрий − матрикс . В матриксе содержатся рибосомы и митохондриальная ДНК (0,5 %), которая имеет кольцевое строение и отвечает за синтез белков митохондрий. Митохондрии имеют все типы РНК (1 %), делятся независимо от деления ядра, в клетке образуются от предсуществующих митохондрий путем деления или почкования. Полупериод жизни митохондрий − 5−10 дней.
Функции . Митохондрии являются центрами энергетической активности клеток. В митохондриях функционируют системы аэробного дыхания и окислительного фосфорелирования. Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТФ-синтетазные комплексы, осуществляющие транспорт электронов и протонов и синтез АТФ. В матриксе располагаются системы окисления ди- и трикарбоновых кислот, ряд систем синтеза липидов, аминокислот и др.
Митохондрии способны передвигаться к местам усиленного потребления энергии. Они могут ассоциировать друг с другом путем тесного сближения или при помощи тяжей. При анаэробном дыхании митохондрии исчезают.
Митохондрии имеют округлую и продолговатую форму диаметром 0,4–0,5 мкм и длиной 1–5 мкм (рис. 1.3).
 |
Количество митохондрий варьирует от единиц до 1 500–2 000 на растительную клетку.
Митохондрии ограничены двумя мембранами: наружной и внутренней, толщина каждой из них 5–6 нм. Наружная мембрана выглядит растянутой, а внутренняя образует складки, называемые гребнями (кристами), различной формы. Пространство между мембранами, в состав которого входит также внутреннее пространство крист, называется межмембранным (перимитохондриальным) пространством. Оно служит средой для внутренней мембраны и матрикса митохондрий.
Митохондрии в целом содержат 65–70 % белка, 25–30 % липидов и небольшое количество нуклеиновых кислот. 70 % от общего содержания липидов составляют фосфолипиды (фосфатидилхолин и фосфатидилэтаноламин). Жирнокислотный состав характеризуется высоким содержанием насыщенных жирных кислот, обеспечивающих «жесткость» мембраны.
В митохондриях локализованы системы аэробного дыхания и окислительного фосфорилирования. В результате дыхания расщепляются органические молекулы, и высвобождается энергия с передачей ее на молекулу АТФ.
Митохондрии содержат белки, РНК, тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеотидах.
Пластиды, наряду с вакуолями и клеточной оболочкой – характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластид различают мембранную систему и более или менее гомогенное вещество – строму. Внутренняя структура хлоропласта довольно сложна. Строма пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами.Тилакоиды собраны в стопки – граны, напоминающие столбики монет.
 |
Хлоропласты, в которых протекает фотосинтез, содержат хлорофиллы и каротиноиды. Размер – 4–5 мкм. В одной клетке мезофилла листа может содержаться 40–50 хлоропластов, в мм2 листа – около 500 000. В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке.
Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых растений и водорослей часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна – это временные хранилища продуктов фотосинтеза. Они могут исчезнуть из хлоропластов, находящихся в темноте всего лишь 24 ч и появиться вновь уже через 3–4 ч после переноса растений на свет.
В изолированных хлоропластах осуществляется синтез РНК, который обычно контролируется только хромосомной ДНК. Образование хлоропластов и синтез находящихся в них пигментов в значительной степени контролируется хромосомной ДНК, малопонятным образом взаимодействующей с ДНК хлоропластов. Тем не менее, в отсутствие собственной ДНК хлоропласты не формируются.
23. Ультраструктура митохондрий, функции
Они участвуют в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
Хромопласты (от греческого сhroma – цвет) – пигментированные пластиды. Многообразные по форме хромопласты не содержат хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и другую окраску. Корнеплоды моркови, плоды томатов окрашены пигментами, которые находятся в хромопластах.
Лейкопласты являются местом накопления запасного вещества – крахмала. Особенно много лейкопластов в клетках клубней картофеля. На свету лейкопласты могут преобразовываться в хлоропласты (клубни картофеля зеленеют). Осенью хлоропласты преобразуются в хромопласты и зеленые листья, и плоды желтеют и краснеют.
Митохондрия – это двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ – источника энергии для жизнедеятельности клетки.
Количество митохондрий в клетках не постоянно, в среднем от нескольких единиц до нескольких тысяч. Там, где процессы синтеза идут интенсивно, их больше. Также варьирует размер митохондрий и их форма (округлые, вытянутые, спиральные, чашевидные и др.). Чаще имеют округлую вытянутую форму, диаметром до 1 микрометра и длиной до 10 мкм. Могут перемещаться в клетке с током цитоплазмы или оставаться в одном положении. Перемещаются к местам, где больше всего требуется выработка энергии.
Следует иметь в виду, что в клетках АТФ синтезируется не только в митохондриях, но и в цитоплазме в процессе гликолиза . Однако эффективность этих реакций невысока. Особенность функции митохондрий в том, что в них протекают реакции не только бескислородного окисления, но и кислородный этап энергетического обмена.
Другими словами, функция митохондрий – активное участие в клеточном дыхании, к которому относят множество реакций окисления органических веществ, переноса протонов водорода и электронов, идущих с выделением энергии, которая аккумулируется в АТФ.
Ферменты митохондрий
Ферменты транслоказы внутренней мембраны митохондрий осуществляют активный транспорт АДФ и АТФ.
В структуре крист выделяют элементарные частицы, состоящие из головки, ножки и основания. На головках, состоящих из фермента АТФазы , происходит синтез АТФ. АТФаза обеспечивает сопряжение фосфорилирования АДФ с реакциями дыхательной цепи.
Компоненты дыхательной цепи находятся в основании элементарных частиц в толще мембраны.
В матриксе находится большая часть ферментов цикла Кребса и окисления жирных кислот.
В результате активности электротранспортной дыхательной цепи ионы водорода поступают в нее из матрикса, а высвобождаются на наружной стороне внутренней мембраны. Это осуществляют определенные мембранные ферменты. Разница в концентрации ионов водорода по разные стороны мембраны приводит к возникновению градиента pH.
Энергию для поддержания градиента поставляет перенос электронов по дыхательной цепи. Иначе ионы водорода диффундировали бы обратно.
Энергия градиента pH используется для синтеза АТФ из АДФ:
АДФ + Ф = АТФ + H 2 O (реакция обратима)
Образующаяся вода ферментативно удаляется. Это, наряду с другими факторами, облегчает протекание реакции слева направо.
Строение и функции ядра растительной клетки.
Ядро – обязательная часть эукариотической клетки. Это место хранения и воспроизведения наследственной информации. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Чаще всего в клетках имеется лишь одно ядро, редко - два или несколько. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью. Снаружи ядро покрыто двойной мембраной – ядерной оболочкой, пронизанной порами (поры ядра - динамичные образования, они могут открываться и закрываться; таким путем может осуществляться регуляция обмена между ядром и цитоплазмой) на краях которых наружная мембрана переходит во внутреннюю. Наружная ядерная мембрана соединяется с мембранными каналами ЭПС. На ней располагаются рибосомы. Внутренняя мембрана может давать впячивания.
Внутреннее содержимое ядра – кариоплазма с погруженными в нее хроматином и ядрышками, и рибосомами. Кариоплазма (нуклеоплазма) представляет собой желеобразный раствор, который заполняет пространство между структурами ядра (хроматином и ядрышками). Она содержит ионы, нуклеотиды, ферменты.
Хроматин – это деспирализованная форма существования хромосом. В деспирализованном состоянии хроматин находится в ядре неделящейся клетке. Хроматин и хромосомы взаимно переходят друг в друга. По химической организации как хроматин, так и хромосомы не отличаются. Химическую основу составляет дезоксирибонуклеопротеин – комплекс ДНК с белками. С помощью белков происходит многоуровневая упаковка молекул ДНК, при этом хроматин приобретает компактную форму.
Ядрышко, обычно шаровидной формы (одно или несколько), не окружено мембраной, содержит фибриллярные белковые нити и РНК. Ядрышки – не постоянные образования, они исчезают в начале деления клетки и восстанавливаются после его окончания. Ядрышки имеются только в неделящихся клетках. В ядрышках происходит формирование рибосом, синтез ядерных белков. Сами же ядрышки образуются на участках вторичных перетяжек хромосом (ядрышковых организаторах).
Ядро – обязательная часть эукариотической клетки. Диаметр ядра колеблется от 5 до 20 мкм. Главная функция ядра – хранение генетического материала в форме ДНК и передача ее дочерним клеткам при клеточном делении. Кроме того, ядро управляет белковыми синтезами, контролирует все процессы жизнедеятельности клетки. (в растительной клетке ядро описал Р.Броун в 1831г., в животной – Т.Шванн в 1838г.).
Химический состав ядра представлен, главным образом, нуклеиновыми кислотами и белками.
Строение и функции митохондрий.
Митохондрии или хондриосомы - «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4-7 мкм и диаметром 0,5-2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена.
Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60-65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК - 1 % и ДНК -0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы.
Митохондрии окружены двойной мембраной. Толщина мембран составляет 6-10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта.
Между мембранами находится заполненное жидкостью перимитохондриальное пространство. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты - кристы в виде пластин и трубочек, они разделяют внутреннее пространство митохондрий на отдельные отсеки. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов).
Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению. Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Рисунок 1. Компоненты мембраны митохондриона. Внешняя мембрана отделяет митохондрию от цитоплазмы. Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии. Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).

Рисунок 2. Мембранные белковые комплексы дыхательной цепи. Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADH2 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Крепления крист и MICOS
Места крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
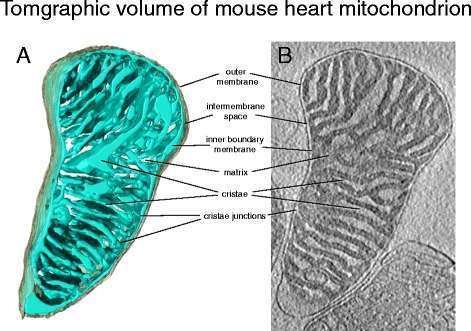
Рисунок 3. Томографический объем митохондрии сердца мыши. А) Трехмерный объем митохондрии сердца мыши, снятый cryo-ET. Наружная мембрана (серая) окутывает внутреннюю мембрану (светло-синяя). Внутренняя мембрана плотно наполнена кристами б) Томографический срез объема. Плотно заполненный матрикс, содержащий большую часть митохондриальных белков, выглядит темным на электронном микроскопе. В то время как межмембранное пространство и просветы крист выглядят светлыми из-за низкой концентрации белков.
Димеры АТФ синтазы
Митохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа . Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами . Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.

Рисунок 4. Двойные ряды АТФ синтазы у семи разных видов.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое. У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.

Рисунок 5. Структура димера АТФ синтазы митохондрии polymella sp. Вид сбоку на V-образный димер АТФ синтазы.
Комплексы и суперкомплексы дыхательной цепи
Протоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду. Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.

Рисунок 6. Комплекс I митохондрии коровьего сердца. Матриксная часть содержит ряд из восьми железно-серных (Fe-S) кластеров, которые направляют электроны из NADH в хинол на пересечении матрикса и мембраны. Мембранная часть состоит из 78 лопастей, включая выкачивающие протоны молекулы.
Комплексы I, III и IV соединяются в суперкомплексы или респирасомы. У пекарских дрожжей (saccharomyces cerevisiae ) нет комплекса I, их суперкомплексы состоят из III и IV. Роль суперкомплексов пока еще не ясна. Предполагают, что это делает транспорт электронов более эффективным, но прямых доказательств этому пока нет.

Рисунок 7. Суперкомплекс митохондрии коровьего сердца. Обратите внимание на дистанцию между комплексами I и III, который надо проделать хинолу. Стрелки – движения электрона в суперкомплексе.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .

Рисунок 8. Ряды димеров АТФ синтазы задают форму кристам. У хребта кристы АФТ синтаза (желтый) образует слив для протонов (красный), протоновые насосы электронной цепи (зеленый) находятся по обоим сторона рядов димеров. Направляя протоны от источника к АТФ синтазе, кристы работают как протоновые направляющие, позволяющие эффективное производство АТФ. Красные стрелки показывают направление потока протонов.
Реорганизация мембраны во время старения
Старение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina , которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.

Рисунок 9. Изменения морфологии внутренней мембраны и димеров АТФ синтазы во время старения митохондрии.
Как видите, полей для будущих исследований очень много. Я предполагаю стык физики и биологии, где физики будут пытаются объяснить почему такая структура более энергетически эффективна. Тем более публикации по квантовой биологии
Митохондрии — это микроскопические мембранные органоиды, которые обеспечивают клетку энергией. Поэтому их называют энергетическими станциями (аккумулятором) клеток.
Митохондрии отсутствуют в клетках простейших организмов, бактерий, энтамеб, которые живут без использования кислорода. Некоторые зеленые водоросли, трипаносомы содержат одну большую митохондрию, а клетки сердечной мышцы, мозга имеют от 100 до 1000 данных органелл.
Особенности строения
Митохондрии относятся к двухмембранным органеллам, имеют внешнюю и внутреннюю оболочки, межмембранное пространство между ними и матрикс.
Внешняя мембрана . Она гладкая, не имеет складок, отграничивает внутреннее содержимое от цитоплазмы. Ширина ее равна 7нм, в составе находятся липиды и белки. Важную роль выполняет порин - белок, образующий каналы во внешней мембране. Они обеспечивают ионный и молекулярный обмен.
Межмембранное пространство . Величина межмембранного пространства около 20нм. Вещество, заполняющее его по составу сходно с цитоплазмой, за исключением крупных молекул, которые могут сюда проникнуть только путем активного транспорта.

Внутренняя мембрана . Построена в основном из белка, только треть отводится на липидные вещества. Большое количество белков являются транспортными, так как внутренняя мембрана лишена свободно проходимых пор. Она формирует много выростов – крист, которые выглядят, как приплюснутые гребни. Окисление органических соединений до CO 2 в митохондриях происходит на мембранах крист. Этот процесс кислородзависимый и осуществляется под действием АТФ-синтетазы. Высвобожденная энергия сохраняется в виде молекул АТФ и используется по мере необходимости.
Матрикс – внутренняя среда митохондрий, имеет зернистую однородную структуру. В электронном микроскопе можно увидеть гранулы и нити в клубках, которые свободно лежат между кристами. В матриксе находится полуавтономная система синтеза белка – здесь расположены ДНК, все виды РНК, рибосомы. Но все же большая часть белков поставляется с ядра, поэтому митохондрии называют полуавтономными органеллами.
Расположение в клетке и деление
Хондриом – это группа митохондрий, которые сосредоточены в одной клетке. Они по-разному располагаются в цитоплазме, что зависит от специализации клеток. Размещение в цитоплазме также зависит от окружающих ее органелл и включений. В клетках растений они занимают периферию, так как к оболочке митохондрии отодвигаются центральной вакуолью. В клетках почечного эпителия мембрана образует выпячивания, между которыми находятся митохондрии.
В стволовых клетках, где энергия используется равномерно всеми органоидами, митохондрии размещены хаотично. В специализированных клетках они, в основном, сосредоточены в местах наибольшего потребления энергии. К примеру, в поперечно-полосатой мускулатуре они расположены возле миофибрилл. В сперматозоидах они спирально охватывают ось жгутика, так как для приведения его в движение и перемещения сперматозоида нужно много энергии. Простейшие, которые передвигаются при помощи ресничек, также содержат большое количество митохондрий у их основания.
Деление . Митохондрии способны к самостоятельному размножению, имея собственный геном. Органеллы делятся с помощью перетяжки или перегородок. Формирование новых митохондрий в разных клетках отличается периодичностью, например, в печеночной ткани они сменяются каждые 10 дней.
Функции в клетке
- Основная функция митохондрий – образование молекул АТФ.
- Депонирование ионов Кальция.
- Участие в обмене воды.
- Синтез предшественников стероидных гормонов.
Молекулярная биология – это наука, изучающая роль митохондрий в метаболизме. В них также идет превращение пирувата в ацетил-коэнзим А, бета-окисление жирных кислот.
| Таблица: строение и функции митохондрий (кратко) | ||
|---|---|---|
| Структурные элементы | Строение | Функции |
| Наружная мембрана | Гладкая оболочка, построена из липидов и белков | Отграничивает внутреннее содержимое от цитоплазмы |
| Межмембранное пространство | Находятся ионы водорода, белки, микромолекулы | Создает протонный градиент |
| Внутренняя мембрана | Образует выпячивания – кристы, содержит белковые транспортные системы | Перенос макромолекул, поддержание протонного градиента |
| Матрикс | Место расположения ферментов цикла Кребса, ДНК, РНК, рибосом | Аэробное окисление с высвобождением энергии, превращение пирувата в ацетил-коэнзим А. |
| Рибосомы | Объединённые две субъединицы | Синтез белка |
Сходство митохондрий и хлоропластов
Общие свойства для митохондрий и хлоропластов обусловлены, прежде всего, наличием двойной мембраны.
Признаки сходства также заключаются в способности самостоятельно синтезировать белок. Эти органеллы имеют свое ДНК, РНК, рибосомы.
И митохондрии и хлоропласты могут делиться с помощью перетяжки.
Объединяет их также возможность продуцировать энергию, митохондрии более специализированы в этой функции, но хлоропласты во время фотосинтезирующих процессов тоже образуют молекулы АТФ. Так, растительные клетки имеют меньше митохондрий, чем животные, потому что частично функции за них выполняют хлоропласты.
Опишем кратко сходства и различия:
- Являются двомембранными органеллами;
- внутренняя мембрана образует выпячивания: для митохондрий характерны кристы, для хлоропластов – тиллакоиды;
- обладают собственным геномом;
- способны синтезировать белки и энергию.
Различаются данные органоиды своими функциями: митохондрии предназначены для синтеза энергии, здесь осуществляется клеточное дыхание, хлоропласты нужны растительным клеткам для фотосинтеза.